
Министерство сельского хозяйства и продовольствия
Российской Федерации
Департамент ветеринарии
Всероссийский научно-исследовательский и технологический
институт биологической промышленности
Утверждаю:
Заместитель начальника
Департамента ветеринарии
В.В.Селиверстов
МЕТОДИЧЕСКИЕ РЕКОМЕНДАЦИИ ПО ИДЕНТИФИКАЦИИ
ПРОИЗВОДСТВЕННЫХ ШТАММОВ ЛЕПТОСПИР И ДИФФЕРЕНЦИАЦИИ
l.interrogans от l.biflexa в реакции ДНК-ДНК-гибридизации
Авторы: В.И.Белоусов. И.Н.Тюрина, С.К.Артюшин
СОКРАЩЕНИЯ, ВСТРЕЧАЮЩИЕСЯ В ТЕКСТЕ РЕКОМЕНДАЦИИ.
т.п.н. - тысяч пар нуклеотидов
гр.С - градусы по Цельсию
10 Х-буфер - десятикратный буфер
20 Х-буфер - двадцатикратный буфер
ДНК - дезоксирибонуклеиновая кислота
sds - додецилсульфат натрия
bsa - бычий сывороточный альбумин
edta - этилендиаминтетраацетат
20 Х ssc - стандартный солевой раствор (3 М nacl, 0,3 М цитрат натрия)
plp - рекомбинантная плазмида
УФ-излучение - ультрафиолетовое излучение
ВВЕДЕНИЕ
Среди болезней общих для человека и животных, весьма су-
щественное значение занимает лептоспироз. Его по праву относят к
числу наиболее распространенных природно-очаговых
зооантропонозов. Род лептоспир по числу известных сероваров
уступает лишь энтеробактериям [5].
В 1983 г. Международный подкомитет по таксономии лептоспир
и спирохет выделил лептоспиры в самостоятельное семейство lep-
tospiraceac в порядке spirochoetalis. В него включен род leptos-
pira, объединяющий l. interrogans - патогенные лептоспиры
и l.biflexa - лептоспиры спирохеты.
Вид патогенных лептоспир включает 212 серовара, которые по
степени антигенного родства объединены в 23 серологические груп-
пы.
Существующая классификация патогенных лептоспир основана на
единственном фенотипическом признаке - антигенной структуре,
изучаемой в перекрестных реакциях иммуноабсорбции.
В последние годы делаются попытки создать классификацию
лептоспир на основании биохимических и генетических критериев
[1,2,3,7,8,9]. Установлено, что ДНК лептоспир относится к at-ти-
пу, а геном лептоспир имеет размер 2000000 пар нуклеотидов
(144). Содержание ГП-пар оснований в ДНК у лептоспир варьирует в
пределах 34,2-41,2 мол % [4].
Хотя вое патогенные лептоспиры объединены в один вид, появ-
ляется все больше данных, указывающих, что этот вид не однороден
и включает в себя ряд группировок.
Лептоспиры по размеру генома (2,0-2,5*10^9 Дальтон) не от-
личаются от других эубактерий, в то же время наблюдается статис-
тически достоверное различие в размере генома между сапрофитными
и паразитическими штаммами лептоспир. При перекрестной
гибридизации ДНК штаммов паразитических лептоспир с ДНК
сапрофитов процент гомологии составляет 3-10% [1,3,7,8,]. Такой
низкий уровень ДНК позволяет рассматривать паразитические и
сапрофитные лептоспиры в качестве самостоятельных видов
генетических комплексов. Анализ подобия нуклеотидных
последовательностей ДНК отдельно у паразитических и сапрофитных
лептоспир показывает, что каждая из этих групп генетически
неоднородна.
Генетический комплекс паразитических лептоспир включает три
группы. Генетический комплекс сапрофитных лептоспир состоит из
четырех групп. Процент гомологии ДНК у штаммов, входящих в гене-
тическую группу, составляет 70-100%, а у штаммов, принадлежащих
к разным генетическим группам, - 20-70% [1]. Таким образом, дан-
ные о степени подобия нуклеотидных последовательностей являются
наиболее информативными.
В настоящее время делаются попытки проводить идентификацию
лептоспир до уровня серовара с применением рестрикционного ана-
лиза их хромосомной ДНК. При специфичном расщеплении ДНК рест-
риктазами образуются фрагменты различной длины, которые
выявляют с помощью электрофореза в агарозном геле. Исследование
полиморфизма длины фрагментов рестрикции ДНК. l. interrogsris
показало, что рестрикционный анализ может быть эффективным
инструментом не только при дифференциации лептоспир, но и при
проведении эпизоотологического надзора.
1. ОБЩИЕ ПОЛОЖЕНИЯ
1.1. Реакция ДНК-ДНК гибридизации является надежным крите-
рием определения родства лептоспир по степени подобия нуклеотид-
ных последовательностей ДНК.
1.2. Методические рекомендации составлены с целью изучения
систематики патогенных лептоопир, поиска возможностей их диффе-
ренциации, идентификации на основе сравнения геномов и использо-
вания в научно-исследовательских учреждениях, ветеринарных лабо-
раториях и биопредприятиях для дифференциации лептоспир вида
l. interrogans от l.biflexa и идентификации производственных
штаммов лептоспир, используемых биологической промышленностью
для изготовления вакцины против лептоспироза животных.
1.3. В реакциях дот-блот гибридизации использовать получен-
ную нами рекомбинантную плазмиду plp 41 о фрагментом ДНК, комп-
лементарным видоспецифическому участку хромосомальной ДНК l.in-
terrogans.
2. ОБОРУДОВАНИЕ
Холодильник бытовой, холодильник низкотемпературный, тер-
мостат, регулируемый на +37°С, 60°С, 55°С, центрифуга фирмы "Эп-
пендорф", трансиллюминатор УФ-излучения, сцинтилляционный счет-
чик радиоактивности, пробирки "Эппендорф", нитроцеллюлозный или
нейлоновый фильтры, аналитические весы, рентгеновская пленка
РМ-1 или РМ-В, рентгеновская кассета с усиливающим экраном.
3. РЕАКТИВЫ, РАСТВОРЫ, МАТЕРИАЛЫ
-0,5 М nacl: растворить 29,22г nacl в 80 мл воды и довести
объем до 100 мл дистиллированной водой.
-20 % sds: растворить 20 г. додецилсульфата натрия в 80 мл
воды, нагреть до 68°С, установить рН 7,2 добавлением нескольких
капель конц. НС1, объем доводят до 100 мл.
-2o x ssc: растворить 175,3 г nacl и 88,2 г цитрата натрия
в 800 мл воды, довести рН до 7,0, добавив несколько капель 10 н
naoh, затем доводят объем до 1 л, разливают на порции.
-бромистый этидный, 10 мг\мл: добавить 1 г бромистого эти-
дия в 100 мл воды, размешать на магнитной мешалке, пока краси-
тель полностью не растворится, хранить в темном месте при 4гр.С.
-буфер ТЕ: 1 мл 1 М трис - НС1, рН 7,4: 200 мкл 0,5 М ЭДТА,
рН 8,0, доводят дистиллированной водой до 100 мл.
-ферменты hind iii, протеиназа К, ДНК полимераза i, ДНК-аза
для мечения ДНК радиоактивным фосфором.
-хлороформ
-фенол: к 10 мл расплавленного фенола добавляют 0,01 г
8-оксихинолина и насыщают равным объемом буфера: сначала 1,0 М
трис-hcl, рН 8,0, а затем 0,1 М трис-hcl, рН 8,0 + 0,2% в-мер-
каптоэтанола до тех пор, пока рН водной фазы не станет больше
7,6. Такой раствор фенола можно хранить 1 мес.
-10Х буфер для ник-трансляции имеет состав: 0,5 М
трис-hcl, рН 7,2; 0,1 М mgcl 2,1 мМ дитиотрейтол и 500 мкг\мл
бычьего сывороточного альбумина. Хранить при 20°С.
-10Х буфер для расщепления ДНК эндонуклеазной hind iii :
10 мМ трио-НС1, рН 7,5; 10 мМ mg-cl 2,1 мМ дитиотрейтол, 50
мМ nаСl.
-агароза и буфер ТВЕ для проведения электрофореза.5Х буфер
Тbe имеет состав: 54 г трис; 27,5 г борная кислота; 2 мл
0,5 m ЭДТА, рН 8,0 доводят дистиллированной водой до 1 литра.
-раствор Денхардта (50x): 5 г фикола, 5 г поливинилпирроли-
дона, 5 г бычьего сывороточного альбумина доводят дистиллирован-
ной водой до 500 мл.
-ДНК спермы лосося
-фрагменты ДНК фага лямбда, полученные при расщеплении pst
i, в качестве маркера молекулярного веса.
-проявитель и закрепитель для фотопленок
-1 М трис: растворить 121,1 г триста в 800 мл воды, довести
до необходимого значения добавлением конц. hcl.
4. ВЫДЕЛЕНИЕ ДНК ИЗ БИОМАССЫ ЛЕПТОСПИР.
5 мл культуры лептоспир центрифугируют 10 мин. при 4
гр.С 10 000 об.\мин. Осадок растворяют в 200 мкл буфера ТЕ.
Затем добавляют 20% sds до конечной концентрации в растворе
0,5% и протеиназы К до конечной концентрации 200 мкг\мл.
Содержимое пробирки оставляют при +37°С на один час. После
инкубирования термостате к раствору добавляют 1\5 его объема
раствор 5 М nacl. далее содержимое пробирки обрабатывают равным
объемом смеси фенол-хлороформ (1:1): тщательно встряхивают и
центрифугируют 5 мин., 14 000 об\мин. Отбирают верхнюю фазу в
чистую пробирку и прибавляют равный объем хлороформа,
тщательно встряхивают и центрифугируют 5 мин., 14 000 об\мин. К
вновь отобранной верхней фазе добавляют 2,5 объема спирта-
ректификата, смесь оставляют при -20°С на 10-16 ч. Затем
центрифугируют 15 мин. при 14 000 об\мин. осадок промывают 70%
спиртом и центрифугируют 10 мин. при 14 000 об\мин. Осадок
сушат в вакуумном эксикаторе и растворяют в 30 мкл буфера ТЕ.
Концентрацию выделенной ДНК в 1 мкл раствора определяют с
помощью электрофореза в 0,8% агарозном геле, сравнивая интенсив-
ность флуоресценции неизвестной и стандартной ДНК.
5. РАДИОАКТИВНОЕ МЕЧЕНИЕ ДНК РЕКОМБИНАНТНОЙ ПЛАЗМИДЫ plp 41
Мечение ДНК plp 41 проводят методом "ник-трансляции".
Для постановки одной реакции берут 0,02-0,05 мкг ДНК, три
немеченых дезоксирибонуклеозидтрифосфата (обычно а-32 Р datp:
3 000 Ки\ммоль, 1 мКи\мл.). Реакцию проводят в объеме 10 мкл
в течение двух часов при +1б°С, внося в пробирку 1 мкл 10 Х
ник-трансляционного буфера, 5 ед. ДНК полимеразы i и фермент
ДНК-азу до конечной концентрации 0,001 мкг\мл. Для очистки
меченной ДНК от невключившихся dntp используют хромотографию на
микроколонках с сефадексом g-50. Процент включения метки рассчитывают с
помощью жидкостного сцинтилляционного счетчика радиоактивности
по соотношению радиоактивности препарата ДНК до хромотографии в
колонке и после нее. Удельную активность меченного зонда
определяют из расчета количества метки (в имп.\мин),
включившейся в 1 мкг ДНК матрицы.
6. ПРОВЕРКА СПЕЦИФИЧНОСТИ ПЛАЗМИДЫ plp 41 НА КОЛЛЕКЦИИ ДНК
ШТАММОВ РАЗЛИЧНЫХ ВИДОВ МИКРООРГАНИЗМОВ
Препараты ДНК различных культур микроорганизмов
(leptospira interrogans, yersinia pseudotuberouiosis,
bordetella bronohlo-septica, e. coli, pasterella irmtocida
(a.b.d), brucella abor-tus, corynebacterium pyogenes,
corynebacterium rinale, mycoplas-ma hyorhinis, mycoplasma
luteus) в объеме 2ОО мкл инкубируют на кипящей водяной бане,
охлаждают на льду и добавляют равный объем 20-кратного ssc
(буфер ssc:3m nacl, 0,3М цитрат натрия).
Предварительно вырезают нитроцеллюлозную или
нейлоновую мембрану и собирают микрофильтрационный аппарат
для олот-блот гибридизации согласно инструкциям фирмы-
изготовителя (bio-rad, richmond, ca). В ячейки аппарата наносят
подготовленные препараты ДНК, после окончания фильтрации
мембрану высушивают на воздухе и прогревают 2 часа при 80 гр.С.
Предгибридизацию проводят Б растворе, содержащем 30-кратный
деионизированный формамид, 5-кратный раствор Денхардта,
двухкратный ssc, 100 мкг\мл спермы лосося, при +47 гр.С 1 час.
К гибридизационному буферу состава: 5-кратный раствор
Денхардта, 2-кратный ssc, 30-кратный формамид, 100 мкг\мл
декстрансульшата добавляют заранее денатурированные кипячением
радиоактивномеченный ДНК-зонд и ДНК спермы лосося (100 мкг\мл).
ДНК-ДНК гибридизацию проводят 18 часов при +47 гр.С.
После инкубации мембрану отмывают от несвязавшейся меченой ДНК
следующим образом: 3 раза по 10 минут в растворе 2-кратного
ssc, 0,2% sds: 2 раза по 15 минут при температуре +6О°С в
растворе 0,2-кратного ssc, 0,2% sds и один раз в течение 10
минут в растворе 0,1-кратного ssc. Отмытую нитроцеллюлозную
мембрану подсушивают и закладывают о рентгеновской пленкой в
кассеты с усиливающим экраном для экспозиции при температуре
-70 грС.
На рисунке 1 показаны радиоавтографы дот-гибридизации
ДНК-зонда plp 41 с препаратами ДНК различных штаммов
микроорганизмов, иммобилизованных на нитроцеллюлозной мембране:
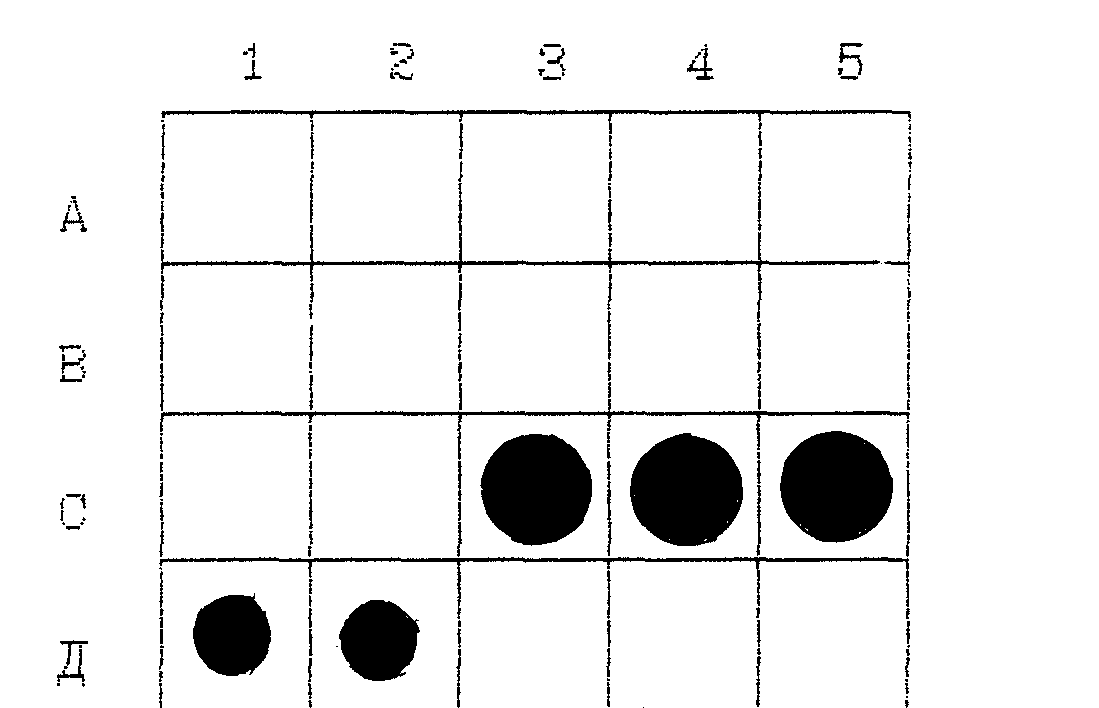 Рис . 1. Радиоавтографы дот-гибридизации ДНК-зонда plp 41
препаратами ДНК разных источников, иммобилизованных на
нитроцеллюлозной мембране:
Линия a:
a1-m.bovis bcg, a2-m.bovis n8,
А3-e.coli, a4-p.multocida,
a5-bordetella bronchioseptica.
Линия В:
b1-y.pseudotuberculosis, b2-m.hyorhinis.
ВЗ-m.leteus, b4-c.pyogenis, b5-c.rinali.
Линия С:
c1-br.abortus, c2-m.hyosi,
c3-l.interrogans серовар pomona шт.ВГНКИ-6,
c4-l.interrogans серовар grippotyphosa
шт. moskva- v., c5- l. interrogans серовар
polonica, шт.493 poland.
Линия d:
d1-плазмида plp 23,
d2-плазмида plp 37.
В местах нанесения ДНК сероваров leptospira interrogans
должны отмечаться положительные сигналы гибридизации в виде тем-
ных пятен на рентгеновской пленке, которые образуются в резуль-
тате засветки связавшимся комплементарно с исследуемыми образца-
ми радиоактивномеченным ДНК-зондом. Микроорганизмы других видов
не должны давать засветки негативной пленки при радиоавтографии.
7. РЕАКЦИЯ Дот-Блот гибридизации
ДЛЯ ДИФФЕРЕНЦИАЦИИ
l.interrogans от l.biflexa
Раствор, содержащий 100-200 мкг ДНК лептоспир, прогревают в
течение 5 мин. при 100°c на водяной бане, резко охлаждают 5 мин.
при 0°С и наносят в виде пятен на предварительно расчерченную на
квадраты нитроцеллюлозную мембрану. После высушивания на воздухе
нитроцеллюлозный фильтр с адсорбировавшейся ДНК инкубируют два
часа при +80 гр.0.
Полученный таким образом фильтр используют для постановки
ДНК-ДНК гибридизации с радиоактивно меченным ДНК-зондом, в ка-
честве которого используется рекомбинантная плазмида plp 41. ДНК
plp 41 метят с помощью реакции "НИК-трансляции". Фильтр с нане-
сенными образцами сначала помещают на один час при 60 гр.С в
предгибридизационную, содержащую 2 Х ssc, 5 Х раствор Денхардта,
100 мкг\мл ДНК спермы лосося, 0,1% sds. Затем фильтр помещают в
гибридизационный раствор, состоящий из тех же компонентов,
что и гибридизационный, и 0,02-0,05 мкг\мл гибридизационного
зонда, меченного радиоактивным фосфором. Фильтр инкубируют при
+ 60°С 18 часов.
Потом фильтр отмывает от несвязавшегося зонда в 0,2 Х ssС с
0,2% sds при комнатной температуре, а далее в 0,1 Х ssc с 0,2%
sds при +60 гр.С, и наконец, в 2 Х ssc при комнатной
температуре.
УЧЕТ РЕЗУЛЬТАТОВ.
Для учета результатов используется радиоавтография. Высушенный
на воздухе при комнатной температуре фильтр закладывают в
рентгеновскую кассету с рентгеновской пленкой и усиливающим
экраном для экспозиции в течение 12-18 ч. при температуре -70
гр.С. Рентгеновскую пленку после экспозиции обрабатывают 10
мин. в растворе проявителя и 10 мин. в растворе закрепителя
(точно также, как это делают при изготовлении фотографий).
Положительный сигнал радиоавтографии свидетельствует, что
нанесенная на данном квадрате нитроцеллюлозной мембраны ДНК от-
носится к l.interrogaris. ДНК, полученная из биомассы
сапрофитных штаммов лептоспир (l.biflexa) не дает засветки
негативной пленки при радиоавтографии.
8. РЕСТРИКЦИЯ ДНК ПРОИЗВОДСТВЕННЫХ ШТАММОВ ЛЕПТОСПИР
Для рестрикции 1 мкг ДНК производственных штаммов лептоспир
используют фермент hind iii.
Рестрикцию проводят в соответствии с рекомендациями Т.Маниатис с
соавт., 1984.[6].
Рестрикционная смесь должна содержать 1 мкг ДНК в объеме 20 мкл.
Добавляют воду к раствору ДНК в стерильной пробирке "Эппендорф"
до объема 18 мкл и перемешивают. Прибавляют 2 мкл буфера 10 л
концентрации для рестрикции (средней ионной силы) и 3-5 ед.
рестриктазы hind iii, тщательно перемешивают и инкубируют смесь
при +37°С в течение 1,5-2,0 ч. Рестрикцию останавливают добавле-
нием 0,5 М ЭДТА, рН 8,0 до конечной концентрации 10мМ. Из каждой
пробирки по 2 мкл и анализируют в геле агарозы, проводя электро-
форез.
9. РЕАКЦИЯ БЛОТ-гибридизации ПО САУЗЕРНУ ДЛЯ
ИДЕНТИФИКАЦИИ ПРОИЗВОДСТВЕННЫХ ШТАММОВ ЛЕПТОСПИР
Метод блот-гибридизации заключается в разделении электрофо-
резом рестрикцированных фрагментов хромосомной ДНК производс-
твенных штаммов лептоспир, в переносе их ДНК на нитроцеллюлозный
или нейлоновый фильтр и последующей гибридизации с находящимся
в омывающем фильтр гибридизационном растворе ДНК-зондом plp 41.
Перенос ДНК на фильтр осуществляют по методике, предложен-
ной Е.southern, 1975. [10].
Препараты ДНК лептоспир после эндонуклеазного расщепления
ферментом hind iii вносят в 0,8% агаразный гель и осуществляют
электрофоретическую разгонку. Окрашивание гелей проводят водным
раствором бромистого этидия (0,5 мкг\мл). Результаты электрофо-
реза наблюдают в проходящем ультрафиолетовом свете на трансиллю-
минаторе и зарисовывают на полиэтиленовой пленке места располо-
жения реотрикцированных фрагментов маркера молекулярной массы. В
качестве молекулярной массы используют фрагменты ДНК фага лямб-
да, полученные при расщеплении pst i.
После завершения электрофореза нуклеиновые кислоты денату-
ририруют в геле, погружая его полностью на 30-40 мин. в две пор-
ции денатурирующего раствора: 1,5 М naoh. Затем нейтрализуют
гель 30-40 мин. двумя порциями нейтрализующего буфера: 1,5
М nacl, 1m трио-hcl, рН 8,0. В ванночку, содержащую 10 Х
буфер ssc, помещают чашку, на которую накладывают сверху
стеклянную пластину, обернутую несколькими слоями пропитанной
10 Х ssc фильтровальной бумаги. Нейтрализованный гель
укладывают нижней стороной вверх на поверхность фильтровальной
бумаги, удаляя воздушные пузырьки. Сверху последовательно
накладывают точно по размеру геля нитроцеллюлозную мембрану
(или нейлоновую), стопку фильтровальной бумаги высотой 10 cм,
стеклянную пластину и груз массой 500 г. Такую систему
оставляют на 12-18 ч. для переноса фрагментов ДНК из геля
на мембрану. После окончания переноса нитроцеллюлозную или
нейлоновую мембрану высушивают на воздухе и прогревают два часа
при 80°С.
Подготовленный таким образом фильтр запаивают в полиэтиле-
новый пакет.
Чтобы блокировать участки неспецифического связывания про-
водят предгибридизацию в течение 1 часа при +65°С в растворе
следующего состава: 2 Х ssc, 5 Х раствор Денхардта, 0,2%.
sds, 100 мкг\мл ДНК спермы лосося. Включение в ДНК радиоактивной
метки осуществляют методом "ник-трансляции". Гибридизацию
ставят при +65 гр.С в течение 18 ч. в растворе, содержащем 2
Х ssc, 5 Х раствор Денхардта, 100 мкг\мл ДНК спермы лосося,
0,25 мкг\мл меченного 32Р ДНК - зонда plp 41.
УЧЕТ РЕЗУЛЬТАТОВ.
Для учета результатов используется радиоавтография.
Высушенный на воздухе при комнатной температуре фильтр зак-
ладывают в рентгеновскую кассету с рентгеновской пленкой и
усиливающим экраном для экспозиции в течение 12-18 ч. при
температуре -70°С. Рентгеновскую пленку после экспозиции
обрабатывают 10 мин. в растворе проявителя и 10 мин. в растворе
закрепителя (точно также, как это делают при изготовлении
фотографий).
С помощью маркера молекулярной массы определяют размер гиб-
ридизующих с ДНК-зондом hind ill-фрагментов геномной ДНК про-
изводственных штаммов лептоспир.
Рис . 1. Радиоавтографы дот-гибридизации ДНК-зонда plp 41
препаратами ДНК разных источников, иммобилизованных на
нитроцеллюлозной мембране:
Линия a:
a1-m.bovis bcg, a2-m.bovis n8,
А3-e.coli, a4-p.multocida,
a5-bordetella bronchioseptica.
Линия В:
b1-y.pseudotuberculosis, b2-m.hyorhinis.
ВЗ-m.leteus, b4-c.pyogenis, b5-c.rinali.
Линия С:
c1-br.abortus, c2-m.hyosi,
c3-l.interrogans серовар pomona шт.ВГНКИ-6,
c4-l.interrogans серовар grippotyphosa
шт. moskva- v., c5- l. interrogans серовар
polonica, шт.493 poland.
Линия d:
d1-плазмида plp 23,
d2-плазмида plp 37.
В местах нанесения ДНК сероваров leptospira interrogans
должны отмечаться положительные сигналы гибридизации в виде тем-
ных пятен на рентгеновской пленке, которые образуются в резуль-
тате засветки связавшимся комплементарно с исследуемыми образца-
ми радиоактивномеченным ДНК-зондом. Микроорганизмы других видов
не должны давать засветки негативной пленки при радиоавтографии.
7. РЕАКЦИЯ Дот-Блот гибридизации
ДЛЯ ДИФФЕРЕНЦИАЦИИ
l.interrogans от l.biflexa
Раствор, содержащий 100-200 мкг ДНК лептоспир, прогревают в
течение 5 мин. при 100°c на водяной бане, резко охлаждают 5 мин.
при 0°С и наносят в виде пятен на предварительно расчерченную на
квадраты нитроцеллюлозную мембрану. После высушивания на воздухе
нитроцеллюлозный фильтр с адсорбировавшейся ДНК инкубируют два
часа при +80 гр.0.
Полученный таким образом фильтр используют для постановки
ДНК-ДНК гибридизации с радиоактивно меченным ДНК-зондом, в ка-
честве которого используется рекомбинантная плазмида plp 41. ДНК
plp 41 метят с помощью реакции "НИК-трансляции". Фильтр с нане-
сенными образцами сначала помещают на один час при 60 гр.С в
предгибридизационную, содержащую 2 Х ssc, 5 Х раствор Денхардта,
100 мкг\мл ДНК спермы лосося, 0,1% sds. Затем фильтр помещают в
гибридизационный раствор, состоящий из тех же компонентов,
что и гибридизационный, и 0,02-0,05 мкг\мл гибридизационного
зонда, меченного радиоактивным фосфором. Фильтр инкубируют при
+ 60°С 18 часов.
Потом фильтр отмывает от несвязавшегося зонда в 0,2 Х ssС с
0,2% sds при комнатной температуре, а далее в 0,1 Х ssc с 0,2%
sds при +60 гр.С, и наконец, в 2 Х ssc при комнатной
температуре.
УЧЕТ РЕЗУЛЬТАТОВ.
Для учета результатов используется радиоавтография. Высушенный
на воздухе при комнатной температуре фильтр закладывают в
рентгеновскую кассету с рентгеновской пленкой и усиливающим
экраном для экспозиции в течение 12-18 ч. при температуре -70
гр.С. Рентгеновскую пленку после экспозиции обрабатывают 10
мин. в растворе проявителя и 10 мин. в растворе закрепителя
(точно также, как это делают при изготовлении фотографий).
Положительный сигнал радиоавтографии свидетельствует, что
нанесенная на данном квадрате нитроцеллюлозной мембраны ДНК от-
носится к l.interrogaris. ДНК, полученная из биомассы
сапрофитных штаммов лептоспир (l.biflexa) не дает засветки
негативной пленки при радиоавтографии.
8. РЕСТРИКЦИЯ ДНК ПРОИЗВОДСТВЕННЫХ ШТАММОВ ЛЕПТОСПИР
Для рестрикции 1 мкг ДНК производственных штаммов лептоспир
используют фермент hind iii.
Рестрикцию проводят в соответствии с рекомендациями Т.Маниатис с
соавт., 1984.[6].
Рестрикционная смесь должна содержать 1 мкг ДНК в объеме 20 мкл.
Добавляют воду к раствору ДНК в стерильной пробирке "Эппендорф"
до объема 18 мкл и перемешивают. Прибавляют 2 мкл буфера 10 л
концентрации для рестрикции (средней ионной силы) и 3-5 ед.
рестриктазы hind iii, тщательно перемешивают и инкубируют смесь
при +37°С в течение 1,5-2,0 ч. Рестрикцию останавливают добавле-
нием 0,5 М ЭДТА, рН 8,0 до конечной концентрации 10мМ. Из каждой
пробирки по 2 мкл и анализируют в геле агарозы, проводя электро-
форез.
9. РЕАКЦИЯ БЛОТ-гибридизации ПО САУЗЕРНУ ДЛЯ
ИДЕНТИФИКАЦИИ ПРОИЗВОДСТВЕННЫХ ШТАММОВ ЛЕПТОСПИР
Метод блот-гибридизации заключается в разделении электрофо-
резом рестрикцированных фрагментов хромосомной ДНК производс-
твенных штаммов лептоспир, в переносе их ДНК на нитроцеллюлозный
или нейлоновый фильтр и последующей гибридизации с находящимся
в омывающем фильтр гибридизационном растворе ДНК-зондом plp 41.
Перенос ДНК на фильтр осуществляют по методике, предложен-
ной Е.southern, 1975. [10].
Препараты ДНК лептоспир после эндонуклеазного расщепления
ферментом hind iii вносят в 0,8% агаразный гель и осуществляют
электрофоретическую разгонку. Окрашивание гелей проводят водным
раствором бромистого этидия (0,5 мкг\мл). Результаты электрофо-
реза наблюдают в проходящем ультрафиолетовом свете на трансиллю-
минаторе и зарисовывают на полиэтиленовой пленке места располо-
жения реотрикцированных фрагментов маркера молекулярной массы. В
качестве молекулярной массы используют фрагменты ДНК фага лямб-
да, полученные при расщеплении pst i.
После завершения электрофореза нуклеиновые кислоты денату-
ририруют в геле, погружая его полностью на 30-40 мин. в две пор-
ции денатурирующего раствора: 1,5 М naoh. Затем нейтрализуют
гель 30-40 мин. двумя порциями нейтрализующего буфера: 1,5
М nacl, 1m трио-hcl, рН 8,0. В ванночку, содержащую 10 Х
буфер ssc, помещают чашку, на которую накладывают сверху
стеклянную пластину, обернутую несколькими слоями пропитанной
10 Х ssc фильтровальной бумаги. Нейтрализованный гель
укладывают нижней стороной вверх на поверхность фильтровальной
бумаги, удаляя воздушные пузырьки. Сверху последовательно
накладывают точно по размеру геля нитроцеллюлозную мембрану
(или нейлоновую), стопку фильтровальной бумаги высотой 10 cм,
стеклянную пластину и груз массой 500 г. Такую систему
оставляют на 12-18 ч. для переноса фрагментов ДНК из геля
на мембрану. После окончания переноса нитроцеллюлозную или
нейлоновую мембрану высушивают на воздухе и прогревают два часа
при 80°С.
Подготовленный таким образом фильтр запаивают в полиэтиле-
новый пакет.
Чтобы блокировать участки неспецифического связывания про-
водят предгибридизацию в течение 1 часа при +65°С в растворе
следующего состава: 2 Х ssc, 5 Х раствор Денхардта, 0,2%.
sds, 100 мкг\мл ДНК спермы лосося. Включение в ДНК радиоактивной
метки осуществляют методом "ник-трансляции". Гибридизацию
ставят при +65 гр.С в течение 18 ч. в растворе, содержащем 2
Х ssc, 5 Х раствор Денхардта, 100 мкг\мл ДНК спермы лосося,
0,25 мкг\мл меченного 32Р ДНК - зонда plp 41.
УЧЕТ РЕЗУЛЬТАТОВ.
Для учета результатов используется радиоавтография.
Высушенный на воздухе при комнатной температуре фильтр зак-
ладывают в рентгеновскую кассету с рентгеновской пленкой и
усиливающим экраном для экспозиции в течение 12-18 ч. при
температуре -70°С. Рентгеновскую пленку после экспозиции
обрабатывают 10 мин. в растворе проявителя и 10 мин. в растворе
закрепителя (точно также, как это делают при изготовлении
фотографий).
С помощью маркера молекулярной массы определяют размер гиб-
ридизующих с ДНК-зондом hind ill-фрагментов геномной ДНК про-
изводственных штаммов лептоспир.
№№
|
шьамм
лептоспир
|
размеры гибридизующихся hind iii-фрагментов
геномной ДНК лептоспир с ДНК-зондом рlр 41
|
1
|
ВГНКИ-1
|
5,1
|
4,7
|
.
|
2,8
|
.
|
1,4
|
2
|
ВГНКИ-2
|
.
|
.
|
4,5
|
3,5
|
.
|
.
|
3
|
ВГНКИ-3
|
5,0
|
.
|
4,5
|
.
|
.
|
.
|
4
|
ВГНКИ-4
|
5,1-2,9
|
.
|
.
|
2,4
|
2,0
|
1,7
|
5
|
ВГНКИ-5
|
.
|
4,8
|
4,5
|
3,5
|
.
|
.
|
6
|
ВГНКИ-6
|
.
|
4,8
|
.
|
3,5
|
.
|
.
|
Производственные штаммы лептоспир отличаются друг от
друга числом и размером гибридизующихся фрагментов, а
также интенсивностью сигнала радиоавтографии (рис 2).
1 2 3 4 5 6 7 8 9 10 11 М
 Рис.2 Радиоавтограф гибридизации с ДНК-зондом plp 41
hind iii - фрагментов хромосомной ДНК штаммов
лептоспир:
1 - ВГНКИ-4 серогруппа tarassovi
2 - ВГНКИ-3 серогруппа canicola
3 - ВГНКИ-5 серогруппа sejroe
4 - ВГНКИ-2 серогруппа icteronaemorrihagiae
5 - ВГНКИ-1 серогруппа grippotyphosa,
6 - ВГНКИ-б серогруппа pomona
7 - moskva v серогруппа grippotyphosa
8 - 493 poland серогруппа
9 - perepelicyn серогруппа tarassovi
10 - Каширский серогруппа canicola
11 - pomona серогруппа pomona
ЛИТЕРАТУРА
1. Артюшин С.k. Изучение генетического родства лептоспир:
Автореферат диссертации кандидата биологических наук. М. 1980,
-15 с.
2. Бернасовская Е.П., Угрюмов Б.П., Вовк А.Д. Лептоспироз.
-Киев, Здоровье, 1989, -152 с.
3.Ежов Г.И., Березова Т.Т., Киктенко В.С. Содержание нуклеиновых кислот и
нуклеотидный состав РНК у некоторых штаммов лептоспир // ЖМЭИ,
1974, n9, с.70-73.
4. Киктенко b.С. Ежов Г.И., Волина Е.Г. Лептоспироз.- М.,
издательство Университета дружбы Народов, 1985, -149с.
5. Малахов Ю.А. Лептоспироз животных. -М: Агропромиздат,
1992, -238с.
6. Маниатис Т., Фрич Э., Сэмбрук Дж. Молекулярное клониро-
вание: Перевод о англ. М: Мир, 1984, -479 с.
7. brendl j.j., rogul m.a. deoxyribonucleic acid hybridiza-
tion among selected leptospiral serotypes // jnt. j of system.
bacteriol. 1974, n24 p. 205-214.
8. haapala d.k., rogul М., evaus l., alexander a. deoxyri-
bonucleic acid base composition and homology studiez of leptos-
pira. // j.bacteriol, 1969, n98, p. 421-428.
9.ramadass p., jarvis b.d.w.,corner r., penny d., marshall r. senetic
characterization of pathogenic leptospira species by dna hybridization // jutern. j of
system bacteriol. 1992, - vol 42, n2 - p.215-219.
10. southern М. detection of specific seguinces amond dna
fragments separated by gel electrophoresis. // j.mol. biol.,
1975, v.95. p. 503-517.
Рис.2 Радиоавтограф гибридизации с ДНК-зондом plp 41
hind iii - фрагментов хромосомной ДНК штаммов
лептоспир:
1 - ВГНКИ-4 серогруппа tarassovi
2 - ВГНКИ-3 серогруппа canicola
3 - ВГНКИ-5 серогруппа sejroe
4 - ВГНКИ-2 серогруппа icteronaemorrihagiae
5 - ВГНКИ-1 серогруппа grippotyphosa,
6 - ВГНКИ-б серогруппа pomona
7 - moskva v серогруппа grippotyphosa
8 - 493 poland серогруппа
9 - perepelicyn серогруппа tarassovi
10 - Каширский серогруппа canicola
11 - pomona серогруппа pomona
ЛИТЕРАТУРА
1. Артюшин С.k. Изучение генетического родства лептоспир:
Автореферат диссертации кандидата биологических наук. М. 1980,
-15 с.
2. Бернасовская Е.П., Угрюмов Б.П., Вовк А.Д. Лептоспироз.
-Киев, Здоровье, 1989, -152 с.
3.Ежов Г.И., Березова Т.Т., Киктенко В.С. Содержание нуклеиновых кислот и
нуклеотидный состав РНК у некоторых штаммов лептоспир // ЖМЭИ,
1974, n9, с.70-73.
4. Киктенко b.С. Ежов Г.И., Волина Е.Г. Лептоспироз.- М.,
издательство Университета дружбы Народов, 1985, -149с.
5. Малахов Ю.А. Лептоспироз животных. -М: Агропромиздат,
1992, -238с.
6. Маниатис Т., Фрич Э., Сэмбрук Дж. Молекулярное клониро-
вание: Перевод о англ. М: Мир, 1984, -479 с.
7. brendl j.j., rogul m.a. deoxyribonucleic acid hybridiza-
tion among selected leptospiral serotypes // jnt. j of system.
bacteriol. 1974, n24 p. 205-214.
8. haapala d.k., rogul М., evaus l., alexander a. deoxyri-
bonucleic acid base composition and homology studiez of leptos-
pira. // j.bacteriol, 1969, n98, p. 421-428.
9.ramadass p., jarvis b.d.w.,corner r., penny d., marshall r. senetic
characterization of pathogenic leptospira species by dna hybridization // jutern. j of
system bacteriol. 1992, - vol 42, n2 - p.215-219.
10. southern М. detection of specific seguinces amond dna
fragments separated by gel electrophoresis. // j.mol. biol.,
1975, v.95. p. 503-517.